Koirarotujen sukupuu paljastaa uusia asioita ja tarkentaa olemassa olevaa tietoa
Monet koirien käyttöominaisuuksiin liittyvät piirteet, kuten paimentaminen ja juoksunopeus, on todennäköisesti kehitetty useammin kuin kerran koiran historian aikana. Vuonna 2017 julkaistussa yhdysvaltalaistutkimuksessa rakentunut koirarotujen sukupuu kertoo rotujen keskinäisestä sukulaisuudesta ja auttaa myös jalostuksessa.
Koiraroduilla on useita erillisiä kehityslinjoja
Koira on ansainnut paikkansa lähes jokaisessa yhteiskunnassa eri puolilla maailmaa. Vuosituhansien aikana koirat ovat avustaneet ihmisiä metsästyksessä ja karjanhoidossa sekä vartioineet taloa ja tiluksia. Koirien käyttöominaisuuksien ja ulkomuodon moninaisuus yhdistettynä koiran ja ihmisen vertaansa vailla olevaan tunnesiteeseen on johtanut yli 350 eri rodun syntyyn.
Koirarotujen muodostumista selvitettiin vuonna 2017 julkaistussa laajassa tutkimuksessa. Elaine Ostranderin tutkimusryhmän uraauurtava tutkimus on Heidi Parkerin kirjoittama (Parker ym. 2017).
Tutkimusaineisto oli tutkimuksen julkaisuhetkellä siihen asti monipuolisin, kun mukana oli 1 346 koiraa 161 rodusta. Näytteitä oli kerätty niin Pohjois-Amerikasta ja Euroopasta kuin Afrikasta ja Aasiastakin. Mukaan oli valikoitu erityisesti rotuja, jotka edustavat nykykoirien ilmiasujen koko kirjoa. Mukana oli myös näytteitä yhdeksästä luonnonvaraisesta koiraeläimestä.
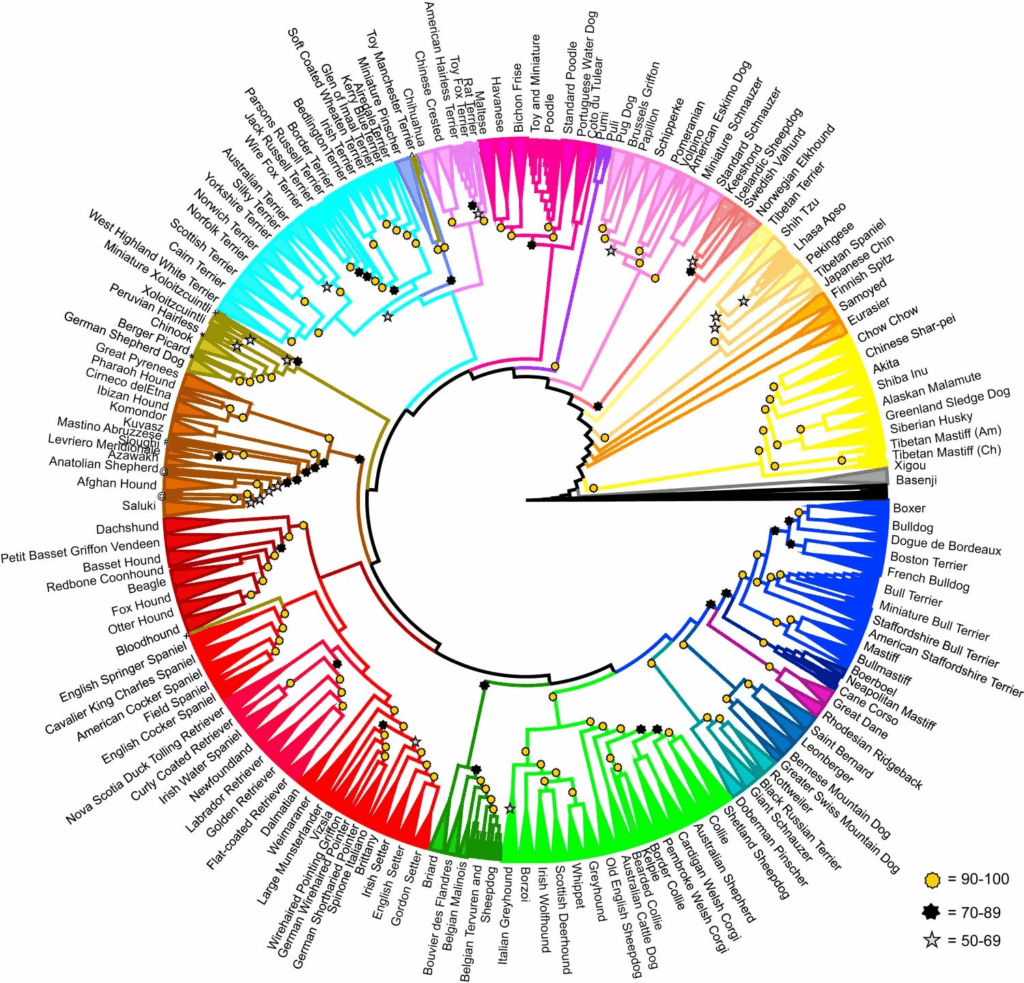
Tutkimuksessa rakentui 161 rodun polveutumishistoriaa kuvaava sukupuu. Sukupuu eli kladogrammi perustuu yksilöiden geneettisen samankaltaisuuden mittaamiseen. Erilliset koirarodut voidaan erottaa sukupuussa omiksi ryhmikseen. Lisäksi sukulaisrodut muodostavat sukupuuhun yhteisiä kehityslinjoja eli kladeja, jotka eroavat muista roduista ja kehityslinjoista. Näitä kehityslinjoja oli 23.
Suurin osa koiraroduista ryhmittyy selvästi omaksi ryhmäkseen, osa on sekalaisempaa
Roduista 146 (91 %) muodosti sukupuuhun selkeät rotukohtaiset ryhmänsä (Kuva 1). Yhdeksän rotua (belgianpaimenkoirat tervueren ja groenendael, cane corso, bullterrieri, kääpiöbullterrieri, rottaterrieri, amerikankarvatonterrieri, lhasa apso ja saluki) muodostivat myös rotukohtaisia ryhmiään, mutta hieman pienemmällä varmuudella.
Neljä rotua (redbone coonhound, sloughi, cane paratore ja jackrussellinterrieri) jakautuivat kehityslinjansa sisällä olevien rotujen kesken, ja kaksi rotua (meksikonkarvatonkoira ja perunkarvatonkoira) jakautuivat erillisten, toisistaan poikkeavien kehityslinjojen kesken.
Osa pienemmällä varmuudella yksiselitteiseksi ryhmäksi asettuneista roduista on rotujen virallisen tunnistamisen osalta melko nuoria tai niillä on ollut pitkään avoimet rotukirjat.
Kahdesta rodusta, cane corsosta ja salukista, kerättiin näytteitä kahdesta maasta, joista toinen oli rodun alkuperämaa. Italian cane corsot muodostavat oman kehityslinjansa, mutta Yhdysvaltojen cane corsot sijoittuvat lähelle napolinmastiffeja. Yhdysvaltojen kannalla on geneettinen yhteys myös muihin mastiffeihin sekä rottweileriin.
Myös yhdysvaltalaiset salukit muodostavat oman kehityslinjansa, kun taas Lähi-idän salukit muodostavat useita ryhmiä sellaisen kehityslinjan sisällä, johon kuuluvat sekä yhdysvaltalaiset salukit että afgaaninvinttikoirat.
Koirien kehityslinjat edustavat varhaisia rotutyyppejä
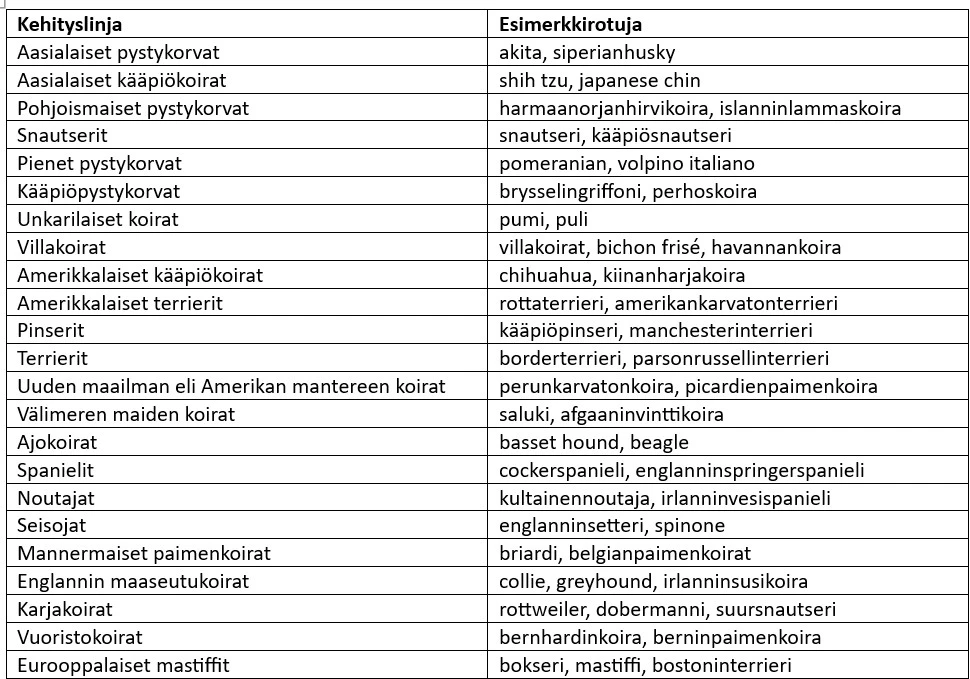
Aineistossa oli havaittavissa 23 erillistä kehityslinjaa, joihin kuului yhteensä 150 rotua eli 93 % aineiston roduista (Taulukko 1). Jokainen kehityslinja koostui 2-18 rodusta. Kehityslinjat edustavat varhaisia rotutyyppejä, jotka olivat olemassa ennen roturekisterien perustamista. Ne heijastavat niihin kuuluvien rotujen yhteistä käyttäytymistä, ulkonäköä ja/tai maantieteellistä alkuperää.
Yksitoista rotua ei ryhmittynyt merkitsevästi mihinkään kehityslinjaan. Näihin rotuihin kuului muun muassa suomenpystykorva. Mahdollisia syitä ryhmittymättömyyteen on kaksi. Aineistossa ei ollut maailman kaikkia koirarotuja, joten tutkimuksessa ei ehkä ollut näytteitä näiden rotujen lähimmistä sukulaisroduista. Toinen syy voi olla, että näissä roduissa on tehty risteytyksiä, jotka eivät ole yhteisiä samankaltaisille roduille.
Taulukko 1. Koirarotujen kehityslinjat (Parker ym. 2017).
Uusi tutkimus tarkensi koirarotujen sukupuuta
Sukupuu kuitenkin tarkentuu, kun aineistoa tulee lisää. Vuonna 2023 julkaistiin kansainvälisen koirien genomihankkeen Dog10k:n tutkimus (Meadows ym. 2023), jossa analysoitiin vielä hieman monipuolisempaa aineistoa eri tavoin.
Aineistossa oli rotukoirien lisäksi näytteitä paikallisista populaatioista ja kyläkoirista useilta eri alueilta. Mukana oli myös luonnonvaraisten koiraeläinten, susien ja kojoottien, näytteitä. Näytteitä oli myös suomalaiskoirista. Kotimaisista roduista mukana olivat suomenpystykorva ja suomenajokoira. Myös pohjanpystykorvasta oli näytteitä.
Nyt sekä suomenpystykorva että pohjanpystykorva ryhmittyivät pystykorvien kehityslinjaan sekä sen alla olevaan pohjoismaisten pystykorvien ryhmään. Suomenajokoira taas ryhmittyi ajokoirien kehityslinjan alaryhmään (Baltian ajokoirat).
Dog10k on vuonna 2016 perustettu maailmanlaajuinen hanke, jonka tavoitteena on tuottaa ja analysoida 10 000 koiraeläimen DNA-sekvenssit ja näin rakentaa kattava tutkimusresurssi. Hankkeeseen osallistuu useita tutkimusryhmiä ja kymmeniä tutkijoita, myös Suomesta.
Koirien kehityslinjojen välillä risteytyksiä
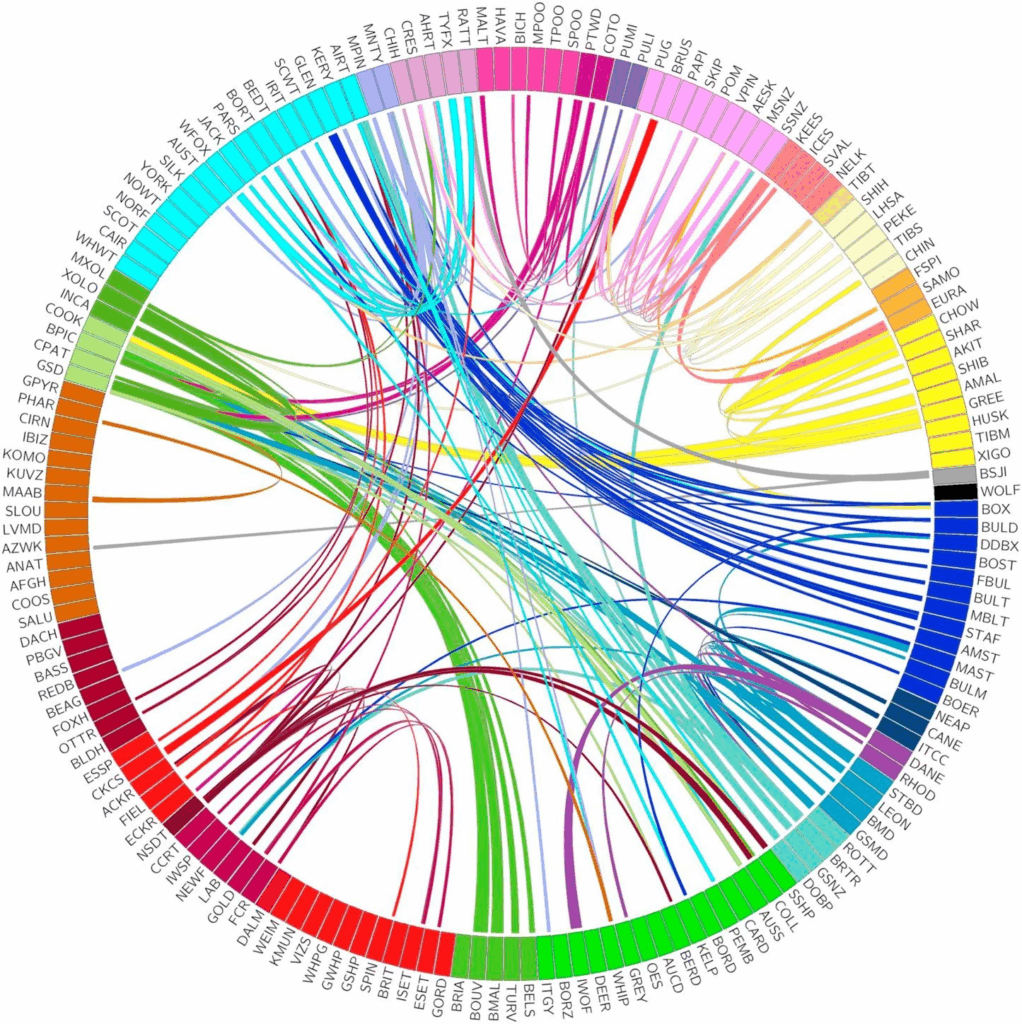
Kehityslinjojen välisen sekoittumisen arvioimiseksi Ostranderin tutkimusryhmä selvitti, kuinka laajasti aineiston koirilla on yhteisiä haplotyyppejä. Haplotyyppi koostuu varianteista, joilla on taipumus periytyä yhdessä. Tietynlainen haplotyyppi kuvastaa kromosomissa lähellä toisiaan sijaitsevien varianttien ainutlaatuista yhdistelmää.
Roduista 117:ssä havaittiin yhteisiä haplotyyppejä muissa kehityslinjoissa olevien koirien kanssa (Kuva 2). Yhteiset haplotyypit ovat osoitus geenivirrasta, jota on tapahtunut roturekisterien syntymisen jälkeisenä aikana, ja joka on johtanut useimpien nykyaikaisten rotujen syntyyn. Kehityslinjojen välillä on siis tehty risteytyksiä uusien ominaisuuksien lisäämiseksi rotuihin.
Noin joka kolmannella näistä 117 rodusta oli yhteys vain yhteen muuhun rotuun kehityslinjansa ulkopuolella. Näin ollen yli puolet roduista, jotka muodostivat tutkimuksen 23 kehityslinjaa, jakavat laajasti haplotyyppejä korkeintaan yhden kehityslinjansa ulkopuolisen rodun kanssa. Nämä rodut näyttävätkin syntyneen pääasiassa alkuperäiseen perustajapopulaatioon perustuvan valinnan kautta sen sijaan, että ne olisi kehitetty rotujen välisillä risteytyksillä.
Ostranderin ryhmän tutkimus havaitsi myös melko tuoreita risteytyksiä. Esimerkiksi eurasier syntyi 1970-luvulla sekoittamalla chow chowta ja muita pystykorvia. Geneettisen sukupuun mukaan eurasier osoitti merkittävää samankaltaisuutta samojedin, keeshondin ja chow chow’n kanssa. Koska kaikki kolme rotua sijaitsivat eri kehityslinjoissa, joilla ei ollut yhteyttä toisiinsa, eurasier sijoittui näiden linjojen väliin ja muodosti oman yhden rodun ryhmänsä.
Joitakin koirarotuja on käytetty usean muun rodun kehittämiseen
Vain kuudella 161 rodusta oli laajoja yhteisiä haplotyyppejä useiden eri rotujen kanssa. Nämä rodut on luotu risteytysten avulla hiljattain tai ne muodostavat suositun nykyrotujen kehittämisessä käytetyn ‘rotukomponentin’.
Suositun rotukomponentin muodostaa esimerkiksi mopsi, joka ryhmittyy kehityslinjassaan tiiviisti eurooppalaisen kääpiökoirarodun, brysselingriffonin läheisyyteen, mutta jolla on yhteisiä haplotyyppejä myös aasialaisten kääpiökoirien kanssa sekä useissa muissa kehityslinjoissa sijaitsevien pienten koirarotujen kanssa. Tämä viittaa todennäköisesti mopsin varhaiseen tuontiin Aasiasta ja sen käyttöön monien pienten rotujen kehittämisessä.
Myös amerikkalaisella rekikoirana käytetyllä chinookilla oli yhteisiä haplotyyppejä laajasti eri kehityslinjojen kanssa. Rotu on luotu hiljattain, ja kirjallisenkin historian mukaan sillä on useita esivanhempia eri roduista.
Yleisesti ottaen risteytyksiä kehityslinjojen välillä on tehty harkitusti ja erityisistä syistä. Niillä on ollut merkittävä rooli monien rotujen kehityksessä.
Koirien kehityslinjojen sisällä risteytyksiä on vähemmän
Useita rotuja sisältävät kehityslinjat voivat muodostua kahdella tavalla. Saman kehityslinjan roduilla on yhteisiä genomialueita, koska ne ovat joko
a) risteytyneet keskenään tai
b) peräisin yhteisistä, muinaisista esivanhemmista, joista ne ovat vähitellen jalostuksen myötä kehittyneet ja eriytyneet omiksi roduikseen.
Asian selvittämiseksi tutkittiin muuttoliikettä 18:ssa neljän tai useamman rodun kehityslinjassa.
Vain Uuden maailman koirien sekä Aasian kääpiökoirien kehityslinjojen sisällä näkyi laajaa geenivaihtoa rotujen välillä. Muiden kehityslinjojen rodut ovat siis todennäköisesti syntyneet muinaisesta rotutyypistä valitsemalla tiettyjä ominaisuuksia, ja niihin on mahdollisesti myös risteytetty muita rotuja kehityslinjan ulkopuolelta ominaisuuksien parantamiseksi.
Aasian kääpiökoiria ei todennäköisesti pidetty erillisinä rotuina, kun niitä vietiin alkuperämaastaan, mikä johti useisiin sekoittumistapahtumiin. Uuden maailman kehityslinjassa havaitut risteymät sen sijaan keskittyvät yksinomaan saksanpaimenkoiriin.
Saksanpaimenkoiran ja italialaisen maatiaiskoiran, cane paratoren (toiselta nimeltään cane toccatore) yhteiset geenit viittaavat todennäköisesti siihen, että näillä roduilla on yhteinen esivanhempi. Saksanpaimenkoira johdettiin 1800-luvun lopulla alkuperältään tuntemattomasta paimenkoirasta.
Yllättävää oli saksanpaimenkoiran haplotyyppien löytyminen kahdelta karvattomalta rodulta (meksikon- ja perunkarvatonkoira). Näitä kahta rotua on ehkä kehitetty saksanpaimenkoiran avulla. Toinen selitys voi olla paimenkoirien yleinen sekoittuminen, kun niitä on viety Etelä-Euroopasta Etelä-Amerikkaan.
Koiria on ollut Amerikan mantereella yli 10 000 vuotta, ja ne ovat todennäköisesti kulkeutuneet sinne Itä-Aasiasta ensimmäisten ihmisten mukana. Aiempien, mitokondriaalisen DNA:n tutkimusten perusteella alkuperäiset Uuden maailman koirat olisivat korvautuneet lähes kokonaan siirtolaiskoirilla, eurooppalaisten kontaktien ja uusien aasialaisten siirtolaisuuksien myötä.
Kun siirtolaiset saapuivat Amerikkaan 1500- ja 1800-luvuilla, he toivat mukanaan karjaa ja siten myös karjan hoitamiseen ja hoivaamiseen tarvittavia koiria. Monet uusista eläimistä syrjäyttivät alkuperäiset, mikä saattaa selittää sen, että Etelä- ja Keski-Amerikan alkuperäisillä karvattomilla roduilla on merkittävä määrä paimenkoiraa genomeissaan, vaikka niitä ei ole jalostettu paimennukseen.
Tutkimuksessa havaittiin, että muinaiset karvattomat rodut ovat risteytyneet laajalti Euroopan paimenkoirien kanssa ja vähäisemmässä määrin myös keskenään.
Tutkimuksessa tunnistettiin myös kaksi Uuden maailman roduista kehittynyttä linjaa, amerikanterrierit ja amerikantoyt. Nämä ovat Pohjois- ja Keski-Amerikasta peräisin olevia pienikokoisia rotuja, joihin kuuluu joukko keskenään sukua olevia terriereitä sekä chihuahua ja kiinanharjakoira.
Kirjallisten tietojen mukaan terrierit juontavat juurensa feistiin, pohjoisamerikkalaiseen maatiaiskoiraan, jota kasvatettiin metsästykseen. Chihuahuan ja kiinanharjakoiran uskotaan olevan peräisin Keski-Amerikasta, vaikka jälkimmäisen nimi viittaakin aasialaiseen syntyperään.
Edellisistä poiketen useimmat Amerikan mantereella kehitetyt uudet rodut ovat syntyneet eurooppalaisten rotujen risteytyksistä ja ryhmittyvät sukupuuhun sen mukaisesti. Esimerkiksi bostoninterrieri ryhmittyy eurooppalaisiin mastiffeihin, kanadalaista alkuperää oleva novascotiannoutaja noutajiin ja Kaliforniassa kehitetty australianpaimenkoira Englannin maaseutukoiriin.
Vanhempien amerikkalaisten rotujen erottuminen sukupuussa huolimatta eurooppalaisesta geenivirrasta viittaa siihen, että näissä roduissa saattaa olla Uuden maailman koirien alkuperäisiä geenejä sekoittuneena eurooppalaisten rotujen geeneihin.
Aivan samoin on tapahtunut eteläamerikkalaisissa ihmispopulaatioissa, joissa on nykyisin sekoittuneena niin alkuperäisväestön kuin eurooppalaisten ja afrikkalaistenkin geenejä. Alkuperäiset Uuden maailman koirien geenit eivät siis ehkä olekaan kokonaan hävinneet nykyisistä koiraroduista, kuten aiemmin on esitetty.
Muuttoliike aiheuttanut myös koirille geneettisiä pullonkauloja
Samoin kuin ihmisillä, myös koirilla maahanmuuttajapopulaatio näyttää olevan vähemmän monimuotoinen kuin populaatio, josta muutetaan.
Kiinasta Yhdysvaltoihin tuodut tiibetinmastiffit ovat muodostuneet geneettisesti itsenäiseksi linjaksi, jossa on havaittavissa myös geneettinen pullonkaula. Pullonkaula näkyy sukusiitoskertoimessa, joka on Kiinan kannassa keskimäärin 0,07 ja Yhdysvaltojen kannassa 0,15.
Sama ilmiö näkyy salukeilla, joiden yhdysvaltalaisen linjan keskimääräinen sukusiitoskerroin on kaksi kertaa suurempi kuin alkuperämaan (0,21 vs. 0,10).
Koirarotujen maantieteelliset kehityslinjat
Maantieteen merkitys rotujen jakautumiseen kehityslinjojen sisällä on mielenkiintoinen. Ostranderin tutkimusryhmän esittämä sukupuu viittaa siihen, että siirtyminen metsästyksestä maanviljelyyn on saattanut käynnistää varhaisen rodunmuodostuksen, ja että tämä tapahtui useilla alueilla itsenäisesti.
Esimerkiksi Englannin maaseudun ja Välimeren alueen kehityslinjoihin kuuluu sekä vinttikoira- että työkoirarotuja, jotka ovat kaksi fyysisesti ja käyttäytymiseltään hyvin erilaista ryhmää. Vinttikoirien fysiikka on rakentunut nopeaan juoksuun, ja niillä on voimakas saalistusvietti. Työkoiriin kuuluvat isot, raskaat laumanvartijat, jotka on jalostettu elämään vartioitavien laumojen keskellä ja toimimaan itsenäisesti ilman vuorovaikutusta ihmisen kanssa. Työkoiriin kuuluvat myös keskikokoiset, ketterät paimenkoirat, jotka on laumanvartijoista poiketen jalostettu toimimaan läheisessä yhteistyössä ihmisen kanssa. Paimenkoirat kontrolloivat ja ohjaavat lauman liikkumista vahingoittamatta paimennettavia eläimiä.
Valinnan kohteena olevista vastakkaisista ominaisuuksista huolimatta vinttikoirat ja työkoirat muodostavat yksittäisen maantieteellistä aluetta kuvaavan geneettisen kehityslinjan niin Englannissa kuin Välimerelläkin. Näiden erillisten kehityslinjojen välillä ei näytä tapahtuneen rotujen muodostamiseen liittyvää sekoittumista, mikä viittaa siihen, että linjojen sisällä olevat rodut ovat kehittyneet eri alueilla itsenäisesti, samoihin ihmisen tarpeisiin. Eri alueilla on tarvittu niin vinttikoiratyyppisiä koiria kuin laumanvartijoita ja paimenkoiriakin, ja koiria on valittu näiden ominaisuuksien perusteella avuksi ihmiselle.
Monet tiettyihin käyttömuotoihin liittyvät piirteet, kuten paimentaminen, juoksunopeus ja suuri koko, on siis todennäköisesti kehitetty useammin kuin kerran nykyaikaisen koiran historian aikana.
Myöskään esimerkiksi Välimeren alueen laumanvartijoiden ja eurooppalaisten mastiffien välillä ei ole geenivirtaa. Näiden rotujen sijoittuminen sukupuussa ja viimeaikaisen sekoittumisen puuttuminen viittaavat siihen, että jättikoko on kehittynyt itsenäisesti eri kehityslinjoissa, ja että se on saattanut olla yksi varhaisimmista ominaisuuksista, joiden perusteella rodut erotettiin toisistaan tuhansia vuosia sitten.
Koirapopulaatiot voidaan siis jakaa geneettisiin ryhmiin myös maantieteen perusteella. Tietyllä alueella olevan koirapopulaation sisällä on jalostettu yleismaailmallisesti merkityksellisiä käytösmalleja ja käyttöominaisuuksia, ja eri alueille on syntynyt samankaltaisia, erillisiä koiratyyppejä jo kauan sitten.
Aineistosta tunnistettiin myös koirarotujen kirjoitetun historian tapahtumia
Tutkijat pystyivät tunnistamaan aineistosta myös rotujen kirjoitettuun historiaan ja rekistereihin kirjattuja tietoja. Aineistosta selvisi esimerkiksi, että kultainennoutaja eriytettiin sileäkarvaisestanoutajasta vuonna 1895. Kirjoitetun historian mukaan kultaisennoutajan historia juontaa juurensa useiden rotujen välisiin risteytyksiin, jotka tapahtuivat vuosina 1868-1890.
Geneettisen aineiston suoma mahdollisuus määrittää risteytysten ajankohtia voi tarkentaa aiempaa tietoa eri rotujen muodostamisesta ja kehityksestä. Kun koiratappelut olivat suosittu viihdemuoto, monia terrieri-, mastiffi- ja bully-rotuja risteytettiin, tarkoituksena tuottaa hyviä koiria kyseiseen lajiin. Käytetyssä tutkimusaineistossa kaikki bull- ja terrieriristeytykset liittyvät Irlannin terrierirotuihin ja ajoittuvat vuosiin 1860-1870. Tämä sopii täydellisesti yhteen historiallisten kuvausten kanssa, joissa kerrotaan koirakilpailujen suosiosta Irlannissa ja kantakirjojen luotettavuuden puutteesta, kun risteytyksiä ei tämän aikakauden aikana dokumentoitu.
Risteymäanalyysin perusteella eniten toisista kehityslinjoista poikkeavat Aasian pystykorvat ja Välimeren maiden koirat, mikä osoittaa, että nämä linjat ovat mahdollisesti vanhempia kuin muut. Tämä sopii hyvin yhteen aiempien tutkimusten kanssa, joiden mukaan varhaisimmat koirat ovat peräisin Keski- ja Itä-Aasiasta.
Näyttää myös siltä, että Aasian pystykorvia on käytetty viimeaikaisissa risteytyksissä, kun taas Välimeren rodut ovat tällä hetkellä enemmän erillisiä.
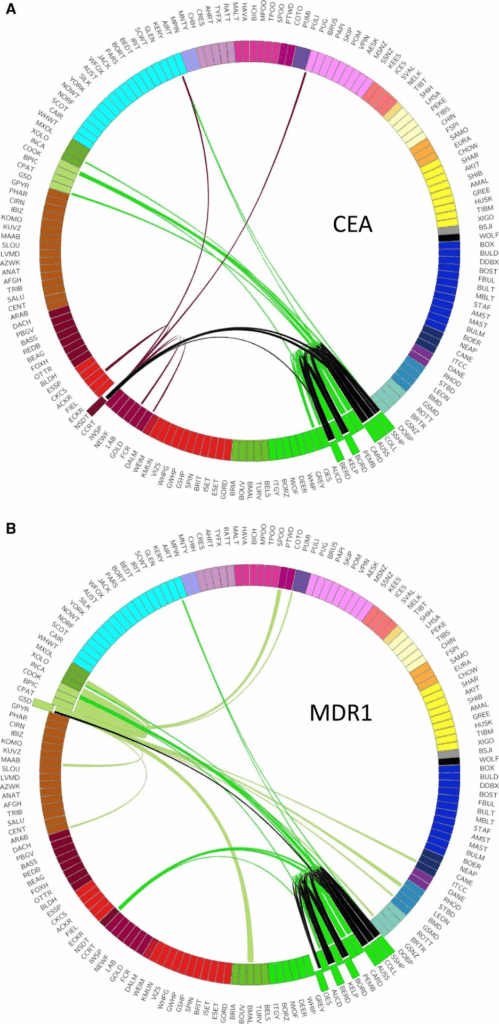
Yhteisiä tautigeenejä yllättävillä koiraroduilla
Risteymäanalyysi antaa tietoa myös kehityslinjojen yhteisistä tautigeeneistä (Kuva 3). Esimerkiksi CEA-silmäsairautta (collie eye anomaly) esiintyy useilla paimenkoirilla, jotka kuuluvat Englannin maaseutukoirien kehityslinjaan, mukaan lukien collie, bordercollie, shetlanninlammaskoira ja australianpaimenkoira. Jo aiemmin on selvitetty, että näillä roduilla näyttää olevan yhteinen esi-isä, jolta sairaus on peräisin.
Sairautta esiintyy kuitenkin myös novascotiannoutajalla, joka on kehitetty Kanadassa tuntemattomasta paikallisten rotujen sekoituksesta. Tämä havainto voitiin nyt selittää, kun analyysi osoitti, että novascotiannoutajalla on yhteisiä haplotyyppejä collien ja/tai shetlanninlammaskoiran kanssa. Näitä rotuja on siis novascotiannoutajan taustalla, ja CEA-geenimuunnoksen alkuperä rodussa juontaa juurensa niihin.
Vastaavasti 10 prosentilla saksanpaimenkoirista on havaittu MDR1-geenin muunnos, joka aiheuttaa hengenvaarallisia lääkeainereaktioita monissa Englannin maaseuturyhmän roduissa. Miksi tämä muunnos on myös saksanpaimenkoiralla, joka ei kuulu tuohon ryhmään? Syy löytyy saksanpaimenkoiran kehittämisajoilta, jolloin rotuun on käytetty australianpaimenkoiraa tai sen edeltäjää. Yhteys ja samalla MDR1-muunnos tulevat saksanpaimenkoirille tätä kautta.
Myös chinookilla on jo aiemmin tunnistettu sama MDR1-geenimuunnos. Parkerin tutkimus paljastikin tämän rodun ja saksanpaimenkoiran sekoittumisen sekä myös aiemmin dokumentoimattoman collieristeytyksen.
MDR1-rotujen kanssa yhteisiä haplotyyppejä löytyy myös meksikonkarvatonkoiralta, joten voidaan ennustaa, että myös tämä harvinainen rotu saattaa kantaa MDR1-geenimuutosta.
Koirarotujen kaksivaiheinen kehitys
Tutkimuksen tulokset tukevat koirarotujen kaksivaiheista kehitystä. Ensin kehittyivät käyttömuodon mukaiset koiratyypit, jotka jalostuivat toiminnallisuuden ja siihen tarvittavan fysiikan mukaan. Tämä näyttäisi tapahtuneen sitä mukaa, kun tarvetta ilmeni ihmisen siirtyessä metsästäjä-keräilijä- ja paimentolaiselämästä maatalouteen ja lopulta kaupunkeihin.
Rotujen prototyypit ovat siis muotoutuneet valikoivien paineiden kautta jo muinaisista ajoista lähtien sen mukaan, mitä työtä koirilta missäkin eniten vaadittiin.
Viimeisten 200 vuoden aikana koiratyyppejä on jaoteltu hyvin spesifisiksi roduiksi jakamalla alkuperäiset, toiminnallisuuden perusteella jalostetut koiratyypit rotuihin, jotka perustuvat pieniin eroihin ulkonäössä. Samalla on saatettu tehdä satunnaisia risteytyksiä ulkomuodon parantamiseksi tai käyttäytymisen muuttamiseksi – esimerkiksi aggressiivisuuden vähentämiseksi ja miellyttämishalun lisäämiseksi.
Koirarotujen sukupuu auttaa roturisteytysten suunnittelussa
Suomessa suunnitellaan risteytyksiä useille roduille. Näiden risteytysten tavoitteena on erityisesti parantaa koirien terveyttä ja tuoda rotuihin niistä kadonneita geenejä. Risteytyskumppanin valinta on joskus työlästä: missä rodussa on ominaisuuksia, joita halutaan, ja mahdollisimman vähän sellaisia, joita ei haluta?
Jos risteytyskumppanina käytetään sukulaisrotua, rodun geenikanta muuttuu maltillisesti, ja jalostus risteytyksen jälkeen on helpompaa.
On myös tärkeää, ettei kumppanilla ole samoja perinnöllisiä sairauksia kuin rodulla, jota risteytyksellä halutaan parantaa.
Ostranderin tutkimusryhmän ja Dog10K-hankkeen tulokset koirarotujen sukupuusta auttavat roturisteytysten suunnittelussa, kun rotujen sukulaisuuksista ja yhteisestä tautiperimästä saadaan koko ajan tarkentuvaa tietoa.
Lisätietoa ja lähteet
Meadows, Kidd, Wang, ym. 2023. Genome sequencing of 2000 canids by the Dog10K consortium advances the understanding of demography, genome function and architecture. Genome Biol 24, 187.
Parker, Dreger, Rimbault ym. 2017. Genomic Analyses Reveal the Influence of Geographic Origin, Migration, and Hybridization on Modern Dog Breed Development. Cell Reports. 19(4): 697-708.
Uutta tietoa lapinkoirien ja hirvikoirien historiasta saatiin tutkimalla DNA:ta 100-200 vuotta vanhoista museoissa säilytetyistä koiran turkeista. Lapinkoirat ja hirvikoirat polveutuvat samasta alkuperäisestä populaatiosta ja ovat sitten eriytyneet omiksi roduikseen. (Wang ym. 2024. Historic dog furs unravel the origin and artificial selection of modern Nordic Lapphund and Elkhound dog breeds. Molecular Biology and Evolution, msae108, https://doi.org/10.1093/molbev/msae108)